

วิวัฒนาการของการรับเสียงในสัตว์มีกระดูกสันหลัง
การรับและวิเคราะห์เสียง จัดเป็นระบบรับสัมผัสที่สำคัญและจำเป็นในสัตว์มีกระดูกสันหลังทุกกลุ่ม แรงผลักดันทางวิวัฒนาการที่สำคัญที่มีผลต่อรูปแบบการรับฟังเสียงมีอยู่ด้วยกัน 2 ประการ ประการแรกคือ การสื่อสาร สัตว์ใช้เสียงเพื่อการสื่อสารในหลายจุดประสงค์ สภาพแวดล้อมจะเป็นตัวกำหนดว่า เสียงแบบใดที่จะถูกส่งผ่านในตัวกลางได้ดี โดยมีการเปลี่ยนแปลงคุณลักษณะน้อยที่สุด ระบบการรับเสียงของสัตว์จะปรับตัวให้เข้ากับข้อจำกัดทางสภาพแวดล้อมเหล่านั้น แรงผลักดันอีกอย่างคือความสามารถของสัตว์ในการบอกทิศทางของแหล่งกำเนิดเสียงที่สนใจได้อย่างถูกต้อง ในสภาพแวดล้อมที่มีเสียงจากหลายแหล่งกำเนิดเสียง มีการกระจายของเสียง และเสียงรบกวน (noise) มากมาย ความสามารถในการแยกเสียงที่สนใจออกจากเสียงอื่น ๆ จัดเป็นปัญหาพื้นฐานของสัตว์ทุกตัวที่จะต้องแก้ไขและเป็นแรงผลักดันทางวิวัฒนาการที่สำคัญยิ่ง
การตรวจจับสัญญาณเพื่อหลีกหนีศัตรูหรือเพื่อล่าเหยื่อเป็นอาหาร จัดเป็นหน้าที่พื้นฐานของระบบการรับเสียง ซึ่งกิจกรรมทั้งสองอย่างนี้ต้องอาศัยทั้งการสื่อสารและความสามารถในการแยกแยะเสียง ด้วยเหตุนี้ โครงสร้างของหูในสัตว์แต่ละกลุ่มจึงมีการปรับเปลี่ยนตลอดเวลาในสายวิวัฒนาการของสัตว์มีกระดูกสันหลัง โดยการเปลี่ยนแปลงที่สำคัญเกิดขึ้นในช่วงรอยต่อของการเปลี่ยนจากการใช้ชีวิตในน้ำขึ้นมาอยู่บนบก ในที่นี้จะกล่าวถึงกลไกการรับและวิเคราะห์เสียงในสัตว์มีกระดูกสันหลังกลุ่มต่าง ๆ ซึ่งสะท้อนให้เห็นถึงการปรับตัวของโครงสร้างหูตลอดช่วงเวลาของวิวัฒนาการ เพื่อให้เกิดประสิทธิภาพในการรับเสียงได้อย่างเหมาะสมในสภาพแวดล้อมที่แตกต่างกัน
1. การรับเสียงในปลา
ในปลา เสียงจากภายนอกสามารถถูกส่งผ่านไปยังหูชั้นในได้หลายทาง เนื่องจากเนื้อเยื่อของปลามีองค์ประกอบหลักส่วนใหญ่เป็นน้ำ และมีสมบัติของคลื่นเสียงคล้ายคลึงกับสภาพแวดล้อมในน้ำที่มันอาศัยอยู่ คลื่นเสียงจึงสามารถผ่านเนื้อเยื่อของปลาเข้าสู่หูชั้นในได้โดยตรง ในปลาบางชนิดมีกระเพาะลม (swim bladder) อยู่ทางด้านบนของท่อทางเดินอาหาร ที่ส่วนปลายของกระเพาะลมมีโครงสร้างยื่นออกมาทางด้านหน้าสองอัน (bulla) ไปติดต่อกับหูชั้นในของแต่ละข้าง ขณะที่บางชนิดกระเพาะลมจะไปเชื่อมต่อกับชุดของกระดูกเล็ก ๆ ที่เรียกว่า Weberian ossicles ซึ่งพัฒนามาจากกระดูกซี่โครง (รูปที่ 1) เมื่อคลื่นเสียงกระทบผ่านลำตัวของปลา จะทำให้เกิดแรงอัดของอากาศในกระเพาะลม แรงสั่นสะเทือนของผนังกระเพาะลม จะถูกส่งผ่าน bulla หรือ Weberian ossicles ไปยังตัวรับเสียงในหูชั้นใน ระบบการทำงานร่วมกันระหว่างกระเพาะลม และ Weberian ossicles มีความคล้ายคลึงกับการทำงานของเยื่อแก้วหูและกระดูกหูสามชิ้นของหูชั้นกลาง (ค้อน ทั่ง และโกลน) ในสัตว์เลี้ยงลูกด้วยน้ำนม (รูปที่ 4)
นอกจากนี้ ปลายังสามารถใช้ระบบเส้นข้างตัว (lateral line) ในการตรวจจับเสียงความถี่ต่ำได้อีกด้วย พบว่าในฉลามและปลากระเบน เส้นข้างตัวมีความไวสูงสุดในการรับเสียงความถี่ช่วง 50 ถึง 150 เฮิรตซ์ ในพวกปลากระดูกแข็งที่มี bulla ยื่นจากกระเพาะลม คลื่นเสียงที่รับโดยเส้นข้างตัวที่บริเวณหัวสามารถส่งผ่านต่อไปยัง bulla แล้วถ่ายทอดไปยังหูชั้นในได้
เนื่องจากหัวของปลามีรูปร่างเรียวแคบ และเสียงเคลื่อนที่ในน้ำได้เร็วกว่าบนบกมาก ทำให้การวิเคราะห์แหล่งกำเนิดเสียงจากความแตกต่างของเวลา หรือความเข้มเสียงที่ไปถึงหูแต่ละข้างเป็นไปได้ยาก ปลาหลายชนิดจึงอาศัยความแตกต่างของขั้วสัญญาณเสียงที่รับโดยเซลล์รับเสียง (hair cells) ในหูชั้นในทั้งสองข้างเป็นตัววิเคราะห์ทิศทางของแหล่งกำเนิดเสียงแทน
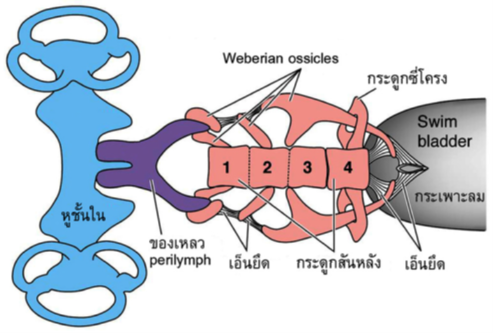
รูปที่ 1การทำงานร่วมกันของกระเพาะลมและ Weberian ossicles ในการส่งผ่านเสียงเข้าสู่หูชั้นใน
สำหรับสัตว์ที่เปลี่ยนวิถีชีวิตจากในน้ำขึ้นมาอยู่บนบก หูที่เคยปรับเข้ากับสภาพแวดล้อมในน้ำก็ต้องเผชิญกับสภาพแวดล้อมใหม่ที่เป็นอากาศแทนเนื่องจากอากาศมีความหนาแน่นต่างจากน้ำมาก ปริมาณของพลังงานเสียงที่ทำให้เกิดการสั่นของโมเลกุลจึงแตกต่างกัน อากาศมีความหนาแน่นน้อยกว่าน้ำ นั่นหมายความว่า อากาศใช้พลังงานในการสั่นของโมเลกุลน้อยกว่าน้ำ เมื่อเสียงในอากาศเคลื่อนที่มาถึงบริเวณผิวรอยต่อระหว่างอากาศและน้ำ พลังงานเสียงจะถูกสะท้อนกลับและไม่สามารถกระตุ้นโมเลกุลในของเหลวได้ ดังนั้น การส่งพลังงานเสียงจากอากาศไปยังของเหลวในหูชั้นใน จึงเป็นปัญหาพื้นฐานของพวกสัตว์สี่เท้า โครงสร้างของหูชั้นนอกและหูชั้นกลางจึงถูกวิวัฒนาการขึ้นเพื่อแก้ไขปัญหานี้ โดยจะช่วยรวบรวม ขยาย และส่งพลังงานเสียงจากอากาศไปยังหูชั้นใน ในปริมาณที่มากพอที่ทำให้เกิดการสั่นของโมเลกุลของของเหลวในหูชั้นใน และกระตุ้นเซลล์รับเสียงได้
2. การรับเสียงในสัตว์สะเทินน้ำสะเทินบก
ในกบหูชั้นนอกประกอบด้วยส่วนของเยื่อแก้วหู หูชั้นกลาง ประกอบด้วย กระดูก columella และกระดูก operculum โดยกระดูก columella จะต่อกับเยื่อแก้วหู พาดผ่านโพรงของหูชั้นกลาง (tympanic cavity) ไปปิดบางส่วนของรูเปิดรูปไข่ (oval window) ส่วนที่เหลือของรูเปิดรูปไข่จะถูกปิดโดยกระดูก operculum ซึ่งจะยึดติดกับกระดูกรองรับขาหน้า (pectoral girdle) โดยกล้ามเนื้อ opercularis เสียงสามารถเข้าถึงหูชั้นในของกบได้ 2 เส้นทางคือ ผ่านทางเยื่อแก้วหู และทาง operculum (รูปที่ 2 ซ้าย) กบจะรับเสียงความถี่ต่ำจากการสั่นสะเทือนของพื้นดินผ่านทางกระดูกรยางค์หน้า และกระดูกรองรับขาหน้าแล้วส่งต่อไปยัง operculum ขณะที่เสียงความถี่สูงกว่าที่มากับอากาศ เช่น เสียงร้องเรียกหาคู่ในฤดูผสมพันธุ์ จะรับผ่านทางเยื่อแก้วหูแล้วส่งต่อไปที่ columella แรงสั่นสะเทือนของเสียงที่ถูกนำผ่านทั้งทางกระดูก columella และ operculum จะถูกส่งผ่านรูเปิดรูปไข่เข้าไปกระตุ้นของเหลวและเซลล์รับเสียงในหูชั้นในต่อไป
ในพวกซาลาแมนเดอร์จะแตกต่างจากกบตรงที่ไม่มีเยื่อแก้วหู ส่วนปลายของกระดูก columella จะต่อติดกับกระดูก squamosal ของกะโหลกผ่านทางเอ็นยึดแทน อย่างไรก็ตามกระดูก columella ยังคงใช้พื้นที่ร่วมกับกระดูก operculum ในการปิดรูเปิดรูปไข่อยู่ เสียงสามารถเดินทางผ่านเข้าหูชั้นในได้ 2 ทางคือ จากกระดูก squamosal ไปยัง columella และจากกระดูกรองรับขาหน้าไปยัง operculum (รูปที่ 2 ขวา) เนื่องจากไม่มีเยื่อแก้วหู ซาลาแมนเดอร์จะมีความไวต่อเสียงในอากาศได้ไม่ดีเท่ากับในกบ
เนื่องจากสัตว์สะเทินน้ำสะเทินบกส่วนใหญ่มีหัวขนาดเล็ก กว้างเพียงไม่กี่เซนติเมตร และรับเสียงได้ดีในช่วงความถี่ที่ไม่สูงมากนัก (ต่ำกว่า 10 กิโลเฮิรตซ์) การวิเคราะห์แหล่งกำเนิดเสียงจากความแตกต่างของเวลาและความเข้มเสียง ระหว่างหูทั้งสองข้างจะใช้ประโยชน์ได้น้อย ในกบจะอาศัยการทำงานร่วมกันระหว่างหูชั้นกลางและช่องปากแทน โดยแรงดันเสียงที่เกิดขึ้นในเยื่อแก้วหูซ้ายและขวาจะถูกส่งผ่านท่อ ยูสเตเชียน (eustachiantube) ไปยังช่องปาก ซึ่งที่นี่จะเกิดการสั่นพ้องของเสียงขึ้น ก่อนส่งไปยังเยื่อแก้วหูของอีกฝั่งหนึ่ง ด้วยวิธีการนี้มันจึงตรวจจับความแตกต่างของเสียงที่มาถึงหูซ้ายและขวาได้ และบอกได้ว่าแหล่งกำเนิดของเสียงนั้นอยู่ในทิศทางใดขณะที่ในซาลาแมนเดอร์ หูชั้นในข้างซ้ายและขวาจะเชื่อมต่อกันผ่านช่องของเหลวที่พาดผ่านกะโหลก ทำให้แรงสั่นสะเทือนของเสียงกระจายจากหูข้างหนึ่งไปยังอีกข้างหนึ่งได้ ช่วยในการวิเคราะห์ความแตกต่างของเสียงระหว่างหูสองข้าง
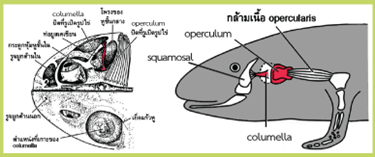
รูปที่ 2 โครงสร้างของหูกบ (ภาพซ้าย) และกลไกการรับเสียงในซาลาแมนเดอร์ (ภาพขวา)
3. การรับเสียงในสัตว์เลื้อยคลานและสัตว์ปีก
โดยทั่วไปหูของสัตว์เลื้อยคลานและสัตว์ปีกจะมีลักษณะใกล้เคียงกัน ท่อรูหูส่วนนอก (external auditory meatus) เริ่มปรากฏครั้งแรกในสัตว์เลื้อยคลานเป็นหลืบตื้น ๆ ส่วนปลายของท่อเป็นที่ตั้งของเยื่อแก้วหูมีกระดูก columella เชื่อมติดกับเยื่อแก้วหู ทอดข้ามโพรงของหูชั้นกลางไปติดกับรูเปิดรูปไข่ (oval window) (รูปที่ 3 ซ้าย) ในสัตว์เลื้อยคลานส่วนใหญ่และสัตว์ปีก เสียงในอากาศจะกระทบเยื่อแก้วหู เกิดการสั่นสะเทือน แล้วส่งต่อไปยังกระดูก columella ผ่านรูเปิดรูปไข่ เข้าสู่หูชั้นในต่อไป อย่างไรก็ตามงูจะไม่เหมือนสัตว์กลุ่มอื่น เพราะไม่มีรูหูส่วนนอกและเยื่อแก้วหูและกระดูก columella จะไปต่อติดกับกระดูก quadrate ซึ่งเชื่อมต่อกับกระดูกขากรรไกรล่างแทน (รูปที่ 3 ขวา) คนส่วนใหญ่มักเข้าใจว่างูนั้นหูหนวก แต่จากงานวิจัยพบว่า มันสามารถตอบสนองต่อเสียงจากการสั่นสะเทือนของพื้นดินและเสียงจากอากาศได้ โดยจะไวต่อเสียงที่มาจากการสั่นสะเทือนของพื้นดินมากกว่า เสียงที่รับจะอยู่ในช่วงความถี่ต่ำและมีความไวในช่วงความถี่แคบ ๆ ขากรรไกรล่างจะรับเสียงที่มากับพื้นดิน แล้วส่งต่อไปยังกระดูก quadrate และcolumella ผ่านรูเปิดรูปไข่ไปยังตัวรับเสียงในหูชั้นใน นักวิทยาศาสตร์เชื่อว่าเยื่อแก้วหูในงูอาจลดรูปหายไป ในช่วงต้นวิวัฒนาการของพวกมันเพื่อปรับตัวสำหรับการใช้ชีวิตในดิน แม้ว่าปัจจุบันนี้ส่วนใหญ่จะอาศัยอยู่บนดิน แต่ก็ยังพบงูโบราณที่ยังคงอาศัยในดินอยู่
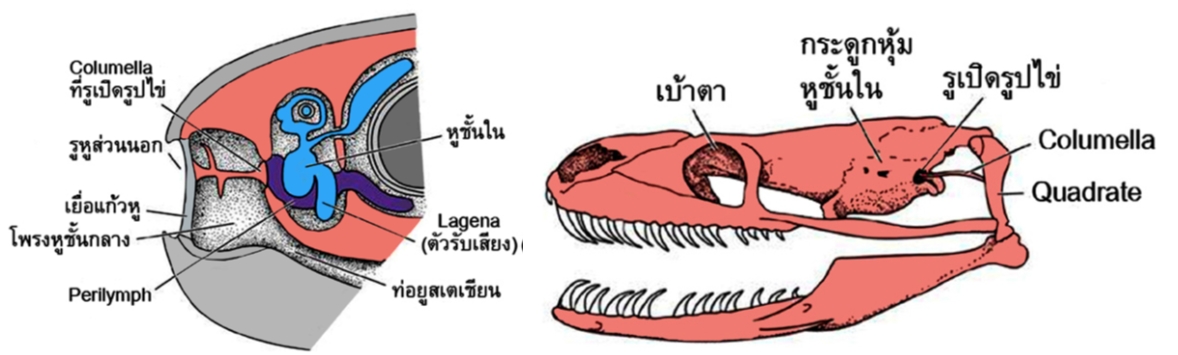
รูปที่ 3 โครงสร้างของหูในสัตว์เลื้อยคลาน (ภาพซ้าย) และกลไกการรับเสียงจากพื้นดินในงู(ภาพขวา)
การวิเคราะห์ทิศทางของเสียงในสัตว์เลื้อยคลานและสัตว์ปีก ยังคงไม่สามารถใช้ประโยชน์จากความแตกต่างของเวลาและความเข้มเสียงระหว่างหูสองข้างได้อย่างเต็มที่ เพราะมีขนาดหัวค่อนข้างแคบ อย่างไรก็ตาม สัตว์ทั้งสองกลุ่มได้พัฒนาวิธีที่ช่วยในการวิเคราะห์ตำแหน่งของเสียงขึ้นหลายวิธี เช่น การที่รูหูทั้งสองข้างมีขนาด รูปร่างหรือตำแหน่งแตกต่างกันหรือการที่สัตว์มีการเคลื่อนที่ของหัวและหูขณะฟังเสียง เป็นต้น
4. การรับเสียงในสัตว์เลี้ยงลูกด้วยน้ำนม
เหตุการณ์สำคัญที่เกิดขึ้น คือการพัฒนาของใบหูที่เคลื่อนไหวได้ (ยกเว้นในกลุ่มที่ออกลูกเป็นไข่ เช่น ตุ่นปากเป็ด) และการปรากฏตัวของกระดูกในหูชั้นกลาง 3 ชิ้น ได้แก่ กระดูกค้อน (malleus) ทั่ง (incus) และโกลน (stapes) (รูปที่ 4) โดยกระดูกโกลนเทียบได้กับกระดูก columella ในสัตว์สี่ขากลุ่มอื่น กระดูกทั้งสามชิ้นจะเรียงตัวต่อติดกันจากเยื่อแก้วหูตามลำดับ และส่วนแผ่นฐานของกระดูกโกลนจะไปประกบติดกับรูเปิดรูปไข่ (oval window) ใบหูจะทำหน้าที่หักเหเสียงเข้าสู่รูหูไปกระทบกับเยื่อแก้วหู กระดูกหูทั้งสามชิ้นจะช่วยขยายแรงสั่นสะเทือนจากเยื่อแก้วหู แล้วถ่ายทอดผ่านเข้าไปในรูเปิดรูปไข่ ทำให้ของเหลวในหูชั้นในสั่นสะเทือน เกิดการกระตุ้นเซลล์รับเสียงได้
การปรากฏของกระดูกหูสามชิ้นนี้ ทำให้สัตว์เลี้ยงลูกด้วยน้ำนมมีประสิทธิภาพในการถ่ายทอดเสียงความถี่สูงสู่หูชั้นใน ได้ดีกว่าการมีกระดูก columella เพียงชิ้นเดียว เนื่องจากปกติกระดูก columella จะมีบางส่วนที่ยังเป็นกระดูกอ่อนเหลืออยู่ ทำให้เกิดการงอและสูญเสียพลังงานได้ระหว่างถ่ายทอดพลังงานไปยังหูชั้นใน ส่วนในสัตว์เลี้ยงลูกด้วยน้ำนม กระดูกทั้งสามชิ้นจะเป็นกระดูกอย่างสมบูรณ์ จึงไม่เกิดการงอและสูญเสียพลังงานน้อยกว่า ด้วยเหตุนี้ จึงไม่แปลกใจว่า ทำไมสัตว์เลี้ยงลูกด้วยน้ำนมถึงสามารถฟังเสียงในช่วงความถี่ที่สูงกว่าในสัตว์มีกระดูกสันหลังกลุ่มอื่น
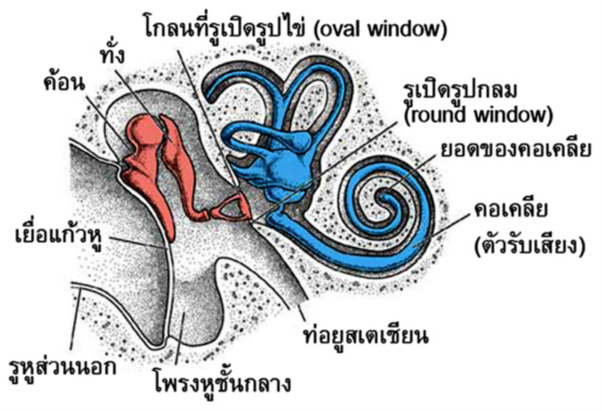
รูปที่ 4 โครงสร้างของหูในสัตว์เลี้ยงลูกด้วยน้ำนม ซึ่งพบกระดูกค้อน ทั่ง และโกลนในหูชั้นกลาง
สำหรับการวิเคราะห์ตำแหน่งของแหล่งกำเนิดเสียง สัตว์เลี้ยงลูกด้วยน้ำนมที่มีหัวขนาดใหญ่สามารถประเมินทิศทางของเสียงได้จากความแตกต่างของเวลาที่เสียงมาถึงรูหูแต่ละข้าง ส่วนในสัตว์ที่มีหัวขนาดเล็ก เช่น หนูหรือค้างคาวบางชนิด จะวิเคราะห์ ทิศทางของเสียงจากความเข้มเสียงที่แตกต่างกันระหว่างหูสองข้าง เนื่องจากเสียงที่มีความถี่ต่ำส่วนใหญ่มักจะเคลื่อนผ่านหัวได้โดยไม่มีการเปลี่ยนแปลง ขณะที่เสียงความถี่สูงสามารถถูกหัวกั้นไว้ได้ ทำให้ความเข้มเสียงที่มาถึงหูทั้งสองข้างไม่เท่ากัน ดังนั้นพวกสัตว์ขนาดเล็กจึงมีวิวัฒนาการเพื่อฟังเสียงที่ความถี่สูงกว่าเพื่อที่จะสามารถใช้ประโยชน์จากความแตกต่างของความเข้มเสียงได้ นอกจากนี้ สัตว์เลี้ยงลูกด้วยน้ำนมยังใช้ประโยชน์ จากการเคลื่อนที่ของหัวและใบหูช่วยในการบอกทิศทางของเสียงอีกด้วย ซึ่งประโยชน์จากใบหูนี้เป็นลักษณะเฉพาะในสัตว์เลี้ยงลูกด้วยน้ำนม
จากที่กล่าวมาทั้งหมดข้างต้น จะเห็นว่า กลไกการรับและวิเคราะห์เสียงในสัตว์มีกระดูกสันหลังนั้น มีรูปแบบที่หลากหลายและซับซ้อนมาก อาศัยการทำงานร่วมกันระหว่างหูกับหลายส่วนของร่างกาย เช่น ช่องปาก ขากรรไกร หรือสมอง และมีการเปลี่ยนแปลงตลอดเวลานับตั้งแต่ยุคเริ่มต้นวิวัฒนาการของสัตว์มีกระดูกสันหลัง สัตว์แต่ละกลุ่มมีการปรับตัวที่แตกต่างกันไป เพื่อที่จะใช้ประโยชน์จากคุณสมบัติของเสียงในสภาพแวดล้อมได้อย่างเหมาะสม เพื่อความอยู่รอดและการดำรงเผ่าพันธุ์ต่อไป
บรรณานุกรม
Fay, R.R., & Popper, A.N. (2000). Evolution of hearing in vertebrates : the inner ears and processing. Hearing Res, 149 : (1-2), 1-10.
Heffner, R.S. (2004). Primate hearing from a mammalian perspective. Anat Rec Part A, 281A : ,1111-1122.
Kardong, K.V. (2006). Vertebrates : Comparative anatomy, function, evolution. Michigan : McGraw-Hill.
Liem, K.F., Bemis, W.E., Walker, W.F., & Grande, L.(2001). Functional anatomy of the vertebrates : an evolutionary perspective. (3rd ed.). Belmont : Thomson Learning.
Manley, G.A. (2010). An evolutionary perspective on middle ears. Hearing Res, 263, 3-8.
McCormick, C.A. (1999). Anatomy of the central auditory pathways of fishes and amphibians. In : Fay. Comparative hearing : fish and amphibians. 155 – 215.
Nummela, S., Thewissen, J.G.M., Bajpai, S., Hussain, T., & Kumar, K., (2007). Sound transmission in archaic and modern whales : anatomical adaptations for underwater hearing. Anat Rec, 290, 716-733.
