

Central dogma
บทนำ
Central Dogma
เป็นแนวคิดอธิบายหน้าที่หลักของดีเอ็นเอ โดยมีเนื้อความว่า ในสิ่งมีชีวิตส่วนใหญ่แล้วข้อมูลทางพันธุกรรมจะถูกเก็บไว้ในรูปของดีเอ็นเอหรือเรียกว่ายีน ดีเอ็นเอเป็นสารพันธุกรรมหลักในการผ่านข้อมูลทางพันธุกรรมหรือถอดรหัสไปสู่อาร์เอ็นเอ (Transcription) แล้วจึงมีการแปลรหัสจากอาร์เอ็นเอสู่โปรตีน (Translation) เพื่อให้เกิดการดำรงชีวิต
ได้มีการศึกษาค้นพบข้อยกเว้นของ Central Dogma คือ พบว่ามีการส่งผ่านข้อมูลทางพันธุกรรมจาก RNA กลับไปสู่ DNA ก่อนแล้วจึงจะถอดและแปลรหัสผ่าน RNA และโปรตีนต่อไป เช่น Retrovirus เป็นไวรัสที่มีอาร์เอ็นเอเป็นสารพันธุกรรม เมื่อไวรัสนี้เข้าไปในเซลล์เจ้าบ้านและเข้าไปจำลอง RNA โดยถอดรหัสจาก RNA ไปเป็น DNA ก่อนโดยใช้เอนไซม์ Reverse transcriptase เมื่อ DNA เข้าสอดแทรกในโครโมโซมของเซลล์เจ้าบ้านก็จะลอกรหัส DNA (DNA replication) ของตนเองไปพร้อมๆกับ DNA ของเซลล์เจ้าบ้าน
กลับไปที่เนื้อหา
 องค์ประกอบและโครงสร้างของ DNA
องค์ประกอบและโครงสร้างของ DNA
การศึกษาโครงสร้างทางเคมีของดีเอ็นเออาศัยข้อมูลที่ได้จากแต่ละวิธีศึกษาประกอบกัน ทำให้มองเห็นโครงสร้างและองค์ประกอบของดีเอ็นเอดีขึ้น หลักฐานเหล่านี้ได้แก่
- ภาพถ่ายของดีเอ็นเอ จากกล้องจุลทรรศน์อิเล็กตรอน ทำให้ทราบว่าโมเลกุลของดีเอ็นเอเป็นสายยาว แต่ไม่สามารถมองเห็นองค์ประกอบทางเคมี
- การวิเคราะห์ดีเอ็นเอทางเคมี มีการศึกษาที่น่าสนใจดังนี้
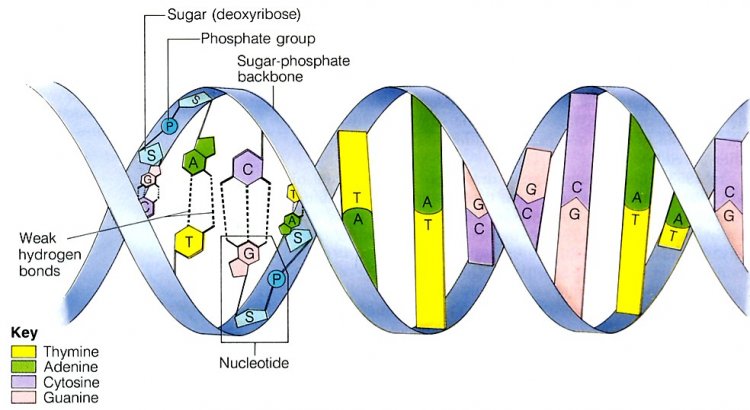
- A. Kossel และ A.T. Levene ศึกษาองค์ประกอบทางเคมีของกรดนิวคลีอิก พบว่า กรดนิวคลีอิกจากเซลล์สัตว์มีธาตุไนโตรเจนเป็นองค์ประกอบหลัก และมีสมบัติเป็นเบส จึงเรียกว่า นิวคลีโอไทด์เบส (Nucleotide base) หรือ ไนโตรจีนัสเบส (Nitrogenous base) ซึ่งมีโครงสร้างทางเคมีแตกต่างกัซึ่งมีโครงสร้างทางเคมีแตกต่างกัน แบ่งเป็น 2 ประเภทคือ
- พิวรีน (Purine) เป็นเบสที่มีโครงสร้างหลักประกอบด้วยวงแหวน 2 วง แบ่งเป็น 2 ชนิดคือ อะดีนีน (Adenine) และกวานีน (Guanine)
- ไพริมิดีน (Pyrimidine) เป็นเบสที่มีโครงสร้างหลักประกอบด้วยวงแหวน 1 วง แบ่งเป็น 2 ชนิดคือ ไทมีน (Thymine) และไซโทซีน (Cytosine)
ต่อมาพบว่ากรดนิวคลีอิกประกอบด้วยน้ำตาลที่มีคาร์บอน 5 อะตอม เรียกว่า น้ำตาลเพนโทส (Pentose sugar) แบ่งเป็น 2 ชนิดคือ น้ำตาลไรโบส (Ribose) เป็นองค์ประกอบของกรดไรโบนิวคลีอิก (Ribonucleic acid : RNA) และน้ำตาลดีออกซีไรโบส (Deoxyribose) เป็นองค์ประกอบของกรดดีออกซีไรบนิวคลีอิก (Deoxyribonucleic acid : DNA) นอกจากนี้ยังพบว่า RNA ต่างจาก DNA ในส่วนของเบสไพริมิดีน โดย RNA จะมีเบสยูราซิล (Uracil) แทนที่เบสไทมีน นอกจากเบสและน้ำตาลเพนโทส ยังมีหมู่ฟอสเฟต เขาจึงเสนอแนวคิดว่า กรดนิวคลีอิกแต่ละโมเลกุลประกอบด้วยหน่วยย่อย (Monomer) เรียกว่า
นิวคลีโอไทด์ (Nucleotide) แต่ละนิวคลีโอไทด์เชื่อมต่อกันเป็นสายยาว โดยมีการสร้างพันธะระหว่างหมู่ฟอสเฟตของนิวคลีโอไทด์หนึ่งกับน้ำตาลของนิวคลีโอไทด์อีกหน่วยหนึ่งที่คาร์บอนตำแหน่ง 3’ เกิดเป็นพอลิเมอร์สายยาวเรียกว่า พอลีนิวคลีโอไทด์ (Polynucleotide)
- Erwin Chargaff วิเคราะห์ปริมาณของนิวคลีโอไทด์ 4 ชนิดในดีเอ็นเอ ทำให้ได้ข้อมูลว่า ดีเอ็นเอในสิ่งมีชีวิตต่างๆ จะมีปริมาณของเบส A = T และ C = G เสมอ เรียกว่า กฎของชาร์กาฟฟ์ (Chargaff’s rules)
- นักเคมีกลุ่มหนึ่งในประเทศอังกฤษ พบว่า นิวคลีโอไทด์แต่ละหน่วยสามารถเชื่อมต่อกันให้เป็นสายพอลีนิวคลีโอไทด์ได้ และสายพอลีนิวคลีโอไทด์แต่ละสายจะแตกต่างกันที่จำนวนนิวคลีโอไทด์ และการจัดเรียงลำดับเบส
- ภาพที่เกิดจากการหักเหของรังสีเอกซ์ผ่าน DNA ในระหว่างปี พ.ศ. 2493 – 2495 M.H.F. Wilkins และ Rosalind Franklin นักฟิสิกส์ชาวอังกฤษ ศึกษาผลึก DNA บริสุทธิ์ด้วยเทคนิคการหักเหของรัวสีเอกซ์ หรือ X – ray diffraction พบว่า โครงสร้างของดีเอ็นเอมีลักษณะเป็นเกลียว (Helix) และประกอบด้วยสายพอลีนิวคลีโอไทด์เกินกว่า 1 สายขึ้นไป และเกลียวแต่ละรอบจะมีระยะทางเท่ากัน
- การเสนอโครงสร้างของดีเอ็นเอ โดย James D. Watson และ Francis Crick มีลักษณะสำคัญโดยสรุปดังนี้
- ประกอบด้วยสายพอลีนิวคลีโอไทด์ 2 สาย เรียงสลับทิศทางกัน คือ ปลายข้าง 3’ ของสายหนึ่งจะประกบกับปลายข้าง 5’ ของอีกสายหนึ่ง
- สายพอลีนิวคลีโอไทด์จะบิดเป็นเกลียวคู่ (Double helix) เวียนขวาหรือหมุนตามเข็มนาฬิกา ดูคล้ายบันไดเวียน มีน้ำตาลดีออกซีไรโบสและหมู่ฟอสเฟตเป็นราวบันได และคู่เบสแต่ละคู่เปรียบเสมือนขั้นบันได
- เกลียวแต่ละรอบจะมีจำนวนคู่เบสเท่ากัน ดังนั้นระยะของเกลียวแต่ละรอบจึงเท่ากัน
- เบส G กับเบส C เกาะกันด้วยพันธะไฮโดรเจน 3 แห่ง ส่วนเบส A กับเบส T เกาะกันด้วยพันธะไฮโดรเจน 2 แห่ง
- ดีเอ็นเอประกอบด้วยพอลีนิวคลีโอไทด์สายคู่ ซึ่งมีความยาวหลายหมื่นคู่เบส การจัดเรียงลำดับเบสที่แตกต่างกันจะทำให้โมเลกุลของดีเอ็นเอมีลักษณะแตกต่างกันมากมาย
กลับไปที่เนื้อหา
การจำลองตัวเองของดีเอ็นเอ (DNA Replication)
ในขณะที่เซลล์มีการแบ่งตัว จะมีการเพิ่มจำนวนโครโมโซมอีก 1 เท่าตัวในระยะอินเตอร์เฟส Sutton เสนอว่า ยีนอยู่บนโครโมโซม James D. Watson และ Francis Crick เสนอว่า เมื่อ DNA ที่เป็นสายเกลียวคู่ จะมีการจำลองตัวเองขึ้นมา สาย Polynucleotide จะแยกออกจากกัน (เหมือนการรูดซิปให้แยกออกจากกัน) โดยพันธะไฮโดรเจนที่ยึดสายทั้งสองสลายตัว เพราะเป็นพันธะที่มีแรงอ่อน เมื่อสาย Polynucleotide ของดีเอ็นเอแยกออกจากกัน ทำให้เบสในแต่ละสายว่างอยู่ ต่อมามีนิวคลีโอไทด์ เข้ามาเกาะเบสที่ว่าง โดยอะดีนีนนิวคลีโอไทด์จะเข้าเกาะกับไทมีนนิวคลีโอไทด์ และไซโทซีนนิวคลีโอไทด์จะเกาะกับกวานีนนิวคลีโอไทด์ ซึ่งยึดกันด้วยพันธะไฮโดรเจน (A จับกับ T และ C จับกับ G เสมอ) นิวคลีโอไทด์ที่มาเกาะจะเชื่อมต่อกันด้วยพันธะฟอสโฟไดเอสเตอร์ และเชื่อมต่อกันเรื่อยๆ จนสิ้นสุดกระบวนการ ทำให้ได้ดีเอ็นเอเกิดขึ้นเป็น 2 โมเลกุล ซึ่งมีลักษณะเหมือนเดิมทุกประการ
ขั้นตอนการสังเคราะห์ดีเอ็นเอจากข้อมูลปัจจุบันมีดังนี้
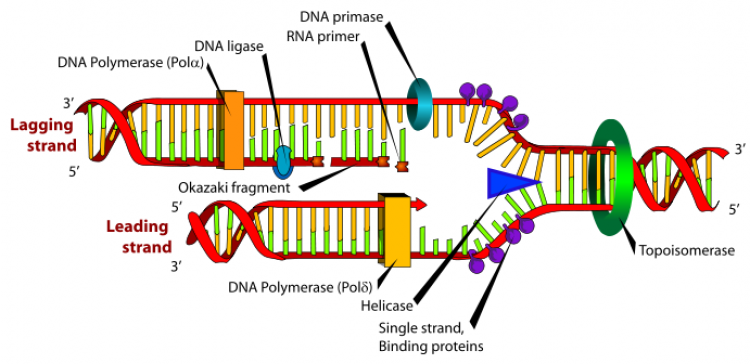
- สาย Polynucleotide แยกตัวออกจากกันโดยพันธะไฮโดรเจนระหว่างคู่เบสสลายตัวด้วยการทำงายของเอนไซม์ Helicase
- Nucleotide อิสระจะเข้าไปจับคู่กับ Nucleotide บนแต่ละสายของ Polynucleotide โดย A จับกับ T , C จับกับ G เสมอโดยทิศทางการสังเคราะห์จะเริ่มจาก 5’ ไป 3’ เสมอ
- Nucleotide ใหม่จะเชื่อมต่อกันเป็นสายยาว เรียกว่า Polynucleotide ในการต่อเชื่อมของ Nucleotide เป็นสาย Polynucleotide ใช้เอนไซม์ DNA polymerase
- Polynucleotide 2 สายจะพันรอบกันและบิดเป็นเกลียวเป็นดีเอ็นเอใหม่ 2 โมเลกุล โดยดีเอ็นเอแต่ละโมเลกุลประกอบด้วย Polynucleotide สายเก่า 1 สายและสายใหม่ 1 สาย เรียกการจำลองดีเอ็นเอแบบนี้ว่าเป็นแบบกึ่งอนุรักษ์ (Semi – conservative replication)
ต่อมา Okazaki พบว่า การสังเคราะห์ DNA เส้นใหม่ขึ้นมาจะต้องมีการสังเคราะห์เป็น 2 แบบ ซึ่งการสร้างสายใหม่มีความยุ่งยาก เนื่องจากการสังเคราะห์ DNA จะเกิดขึ้นในทิศทางจาก 5’ ไป 3’ เท่านั้น การสังเคราะห์ดีเอ็นเอสายหนึ่งเกิดขึ้นได้ไม่ยาก เพราะสายที่เป็นต้นแบบ (Template) มีการแยกตัวในทิศทาง 3’ ไป 5’ จึงสร้างสายใหม่เป็นสายยาวๆ มาเข้าคู่ในทิศทาง 5’ ไป 3’ ได้ทันที สายที่สร้างใหม่ที่มีทิศทางจาก 5’ ไป 3’ นี้เรียกว่า สายนำ (Leading strand) ส่วนการสังเคราะห์อีกสายหนึ่งนั้นสายต้นแบบมีการแยกตัวไปในทิศทางจาก 5’ ไป 3' สายใหม่ที่สร้างขึ้นจึงต้องสร้างทีละช่วงได้เป็นชิ้นส่วนสั้นๆ ประมาณ 1,000 – 2,000 นิวคลีโอไทด์ เรียกว่า Okazaki fragment ก่อน แล้วจึงเชื่อมต่อเป็นสายยาวเส้นเดียวภายหลังเมื่อปลาย 3’ มาอยู่ชิดกับปลาย 5’ ของอีกเส้นที่อยู่ข้างๆ โดยอาศัยเอนไซม์ DNA ligase สายที่สร้างโดยวิธีนี้เรียกว่า สายตาม (Lagging strand)
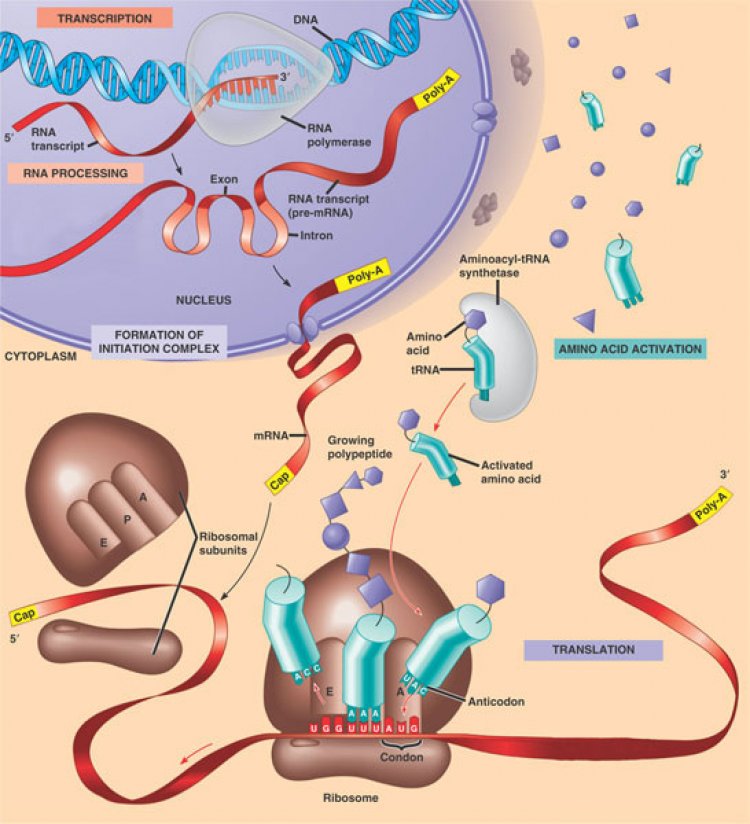
การสังเคราะห์ RNA โดยมี DNA เป็นแม่พิมพ์ (Transcription) มีขั้นตอนดังนี้
- ในบริเวณที่จะมีการสังเคราะห์ RNA Polynucleotide ของ DNA คลายเกลียวแยกออกจากกัน
- นิวคลีโอไทด์ของ RNA จะเข้าจับกับเบสของ DNA โดยเบส A, T, G และ C ของ RNA จะเข้าคู่จับกับเบส T, A, C และ G ของ DNA ตามลำดับ
- การสังเคราะห์ RNA จะมีทิศทางเริ่มจากปลาย 3’ ไปยังปลาย 5’ ของ DNA ดังนั้นโมเลกุลของ RNA ที่สังเคราะห์ได้จะมีทิศทางจาก 5’ ไป 3’
- การสังเคราะห์ RNA นั้นใช้ DNA เพียงสายเดียวเป็นต้นแบบ จึงทำให้ RNA มีโครงสร้างเป็นสายเดียว (Single strand) เรียกว่า ทรานสคริปชัน (Transcription)
- นิวคลีโอไทด์ของ RNA จะเชื่อมต่อกันโดยใช้เอนไซม์ RNA polymerase
จากการศึกษาพบว่า RNA เป็นกรดนิวคลีอิกที่พบทั้งในนิวเคลียส ไซโทพลาสซึม คลอโรพลาสต์และไมโทคอนเดรีย จำแนกออกเป็น 3 กลุ่มคือ
- Messenger RNA (mRNA) เป็น RNA ที่มีขนาดใหญ่ที่สุด มีน้ำหนักโมเลกุลประมาณ 30,000 – 300,000 ดาลตัน ประกอบด้วยนิวคลีโอไทด์ประมาณ 900 – 1,000 นิวคลีโอไทด์ มีปริมาณค่อนข้างน้อยประมาณ 4% ของ RNA ภายในเซลล์ และพบเกาะติดกับไรโบโซมในขณะที่มีการสังเคราะห์โปรตีน เพราะลำดับเบสของ mRNA ทำหน้าที่นำรหัสพันธุกรรม (Genetic code) จาก DNA เพื่อกำหนดชนิดกรดอะมิโนที่ใช้สังเคราะห์โปรตีน
- Transfer RNA (tRNA) เป็น RNA ที่มีขนาดเล็กที่สุด มีน้ำหนักโมเลกุลประมาณ 25,000 ดาลตัน ประกอบด้วยนิวคลีโอไทด์ประมาณ 75 – 80 นิวคลีโอไทด์ มีปริมาณประมาณ 15% ของ RNA ภายในเซลล์ และมีแอนติโคดอน (Anticodon) ซึ่งประกอบด้วยเบส 3 โมเลกุล ที่มีลำดับเข้าคู่กับโคดอน (Codon) ของ mRNA ได้ ทำให้สามารถนำกรดอะมิโนตามรหัสพันธุกรรมมายัง mRNA เพื่อใช้ในการสังเคราะห์โปรตีนได้
Ribosomal RNA (rRNA) เป็น RNA ที่มีปริมาณมากที่สุด พบประมาณ 85% ของ RNA ในเซลล์ พบเป็นองค์ประกอบของไรโบโซมของสิ่งมีชีวิตชนิดต่างๆ จึงมีขนาดโมเลกุลแตกต่างกันขึ้นอยู่กับชนิดของสิ่งมีชีวิตเหล่านั้น rRNA ทำหน้าที่เป็นแหล่งยึดของ mRNA ในขณะที่มีการแปลรหัสเพื่อสังเคราะห์โปรตีน
![]()
กลับไปที่เนื้อหา

 (1781113)
(1781113)  (17503)
(17503) 
