

โครโมโซม
โครมาทิน
โครมาทิน (Chromatin)
ในเซลล์พวกยูคาริโอต (Eukaryotic cell) สารพันธุกรรมคือ DNA อยู่รวมกับโปรตีน และมีการขดตัวเป็นเส้นใยโครมาทิน โดยประกอบด้วย DNA และโปรตีน ประมาณ 1/3 และ 2/3 ตามลำดับ โปรตีนที่พบมี 2 ชนิดคือ ฮิสโทน (Histone) และนอนฮิสโตน (Non - histone) ในระยะอินเตอร์เฟส ส่วนของโครมาทินที่ขดตัวมากเรียกว่า เฮเทอโรโครมาทิน (Heterochromatin) และขดตัวน้อยเรียกว่า ยูโครมาทิน (Euchromatin) สามารถจำแนกโครมาทินออกเป็น 2 ชนิดตามการติดสีเข้มหรือสีจางของโครโมโซม ซึ่งอยู่ในระยะอินเตอร์เฟส ได้แก่
- ยูโครมาทิน (Euchromatin) เป็นโครมาทินที่มีลักษณะยืดยาว ติดสีจาง เส้นใยของโครมาทินมีขนาดเส้นผ่านศูนย์กลางประมาณ 40 – 70 Å ได้มีข้อพิสูจน์ด้วยวิธีการ Autoradiograph ว่าเป็นโครมาทินส่วนที่ดีเอ็นเอมีการทำงานคือ มีการถอดรหัสพันธุกรรม (Transcription) เกิดขึ้น
- เฮเทอโรโครมาทิน (Heterochromatin) เป็นโครมาทินที่หดสั้นมากตลอกระยะเวลาของอินเตอร์เฟสและการแบ่งเซลล์ เส้นใยของโครมาทินชนิดนี้มีขนาดเส้นผ่านศูนย์กลาง 200 – 300 Å DNA ในส่วนของเฮเทอโรโครมาทิน ไม่มีการถอดรหัสพันธุกรรม จากการศึกษาโมเลกุลของดีเอ็นเอ และการศึกษาทางไซโตจินิติค (Cytogenetic) พบว่าเฮเทอโรโครมาทินจำแนกเป็น 2 ชนิดคือ
2.1 Facultative heterochromatin เมื่อเซลล์มีการแปรรูปร่างไปส่วนของเอเทอโรโครมาทินในบางโครโมโซมอาจมีพฤติกรรมเปลี่ยนแปลงไปได้ โดยการหดตัวมากขึ้นและไม่มี Transcription (หยุดการทำงานหรือเรียกว่า inactivation) ตัวอย่างของ Facutative heterochromatin ที่เกิดกับคน คือ inactivation ของโครโมโซม X ในเพศหญิง หญิงปกติจะมีโครโมโซม X 1 คู่ ซึ่ง 1 ใน 2 ของโครโมโซม X จะมี inactivation ตลอดชีวิตของเซลล์นั้น
2.2 Constitutive heterochromatin เป็นโครมาทินที่หดตัวมากและไม่มีการแสดง Transcription เกิดขึ้นตลอดการแปรรูปของเซลล์นั้นๆ ตำแหน่งของ Constitutive heterochromatin บนโครโมโซมทำได้โดยการย้อมสีชนิด C – band ตำแหน่งที่พบบนโครโมโซมคน คือ บริเวณเซนโทรเมียร์ของทุกๆ โครโมโซม ยกเว้นโครโมโซม Y ที่พบบริเวณปลายแขนข้างยาวของ Y
กลับไปที่เนื้อหา
โครโมโซม (Chromosome)
โครโมโซมหรือนิวคลีออยด์ (Nucleoid) ของสิ่งมีชีวิตกลุ่ม Prokaryote จะมีองค์ประกอบง่ายๆ ส่วนใหญ่เป็นดีเอ็นเอเกลียวคู่อยู่ร่วมกับโปรตีนจำนวนน้อย ซึ่งดีเอ็นเอนี้มีความยาวมากกว่าเส้นผ่านศูนย์กลางของเซลล์ มีลักษณะเป็นวงแหวน แต่ละวงของโครโมโซมจะมีจุดเริ่มต้นการจำลองตัวเองเพียงจุดเดียว
ดีเอ็นเอพันขดกันในลักษณะของการพันเกลียวซ้อนเกลียว (Supercoli) สำหรับโครโมโซมของสิ่งมีชีวิตกลุ่ม Eukaryote มีความซับซ้อนมากกว่า
ความสัมพันธ์ของดีเอ็นเอกับนิวคลีโอโซม
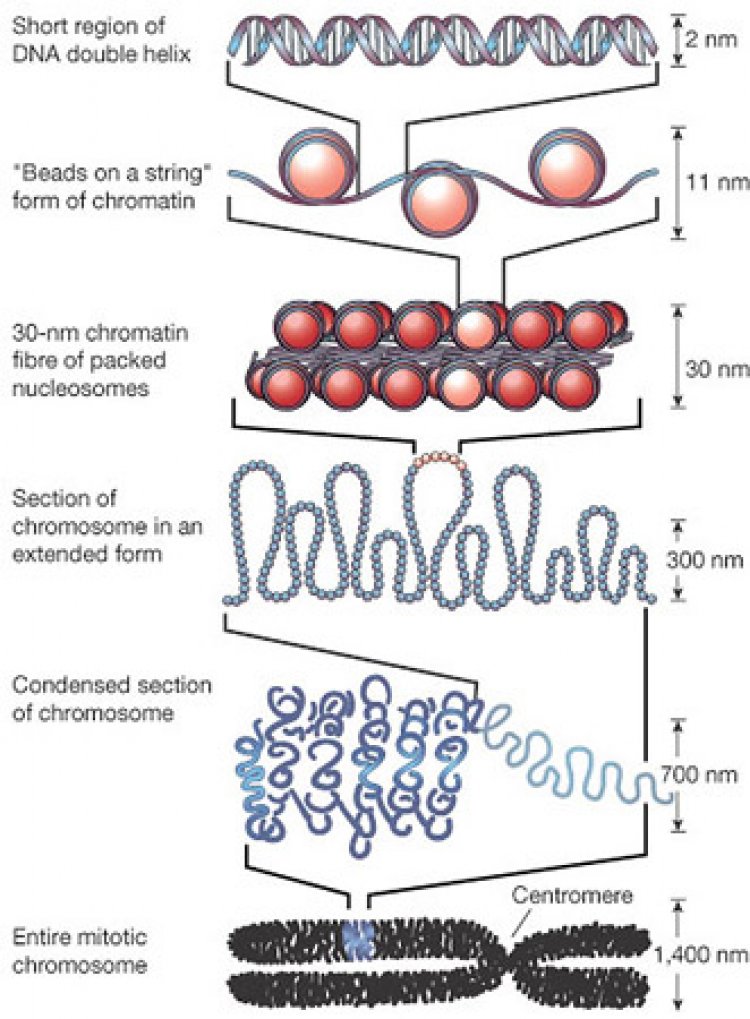
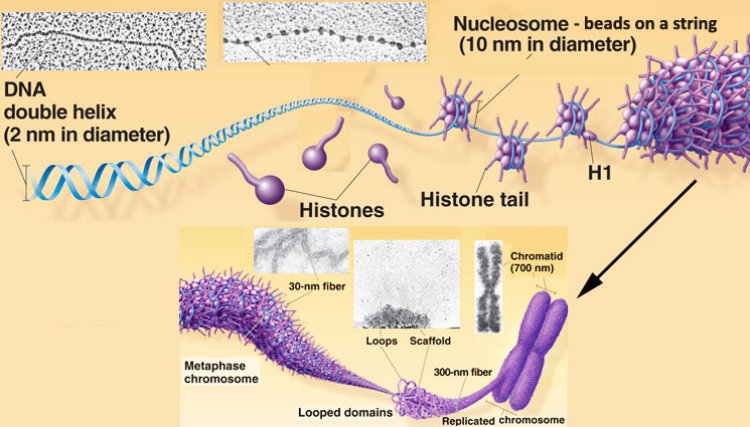
ดีเอ็นเอจัดเป็นสารพันธุกรรมสากลของสิ่งมีชีวิต ประกอบด้วยนิวคลีโอไทด์ 4 ชนิด ได้แก่ dAMP, dCMP, dGMP และ dTMP จำนวนต่างๆกันประกอบเป็นสายดีเอ็นเอ ในโมเลกุลของดีเอ็นเอ 1 โมเลกุลมีสายดีเอ็นเอ 2 สาย ทิศทางตรงกันข้ามจับกันด้วยพันธะไฮโดรเจนระหว่างเบสคู่สม โดยพันเป็นเกลียวหมุนเวียนขวา (Double helix) และพันรอบโปรตีนฮิสโตน (Histone) H2A, H2B, H3 และ H4 อย่างละ 2 โมเลกุลที่จับกันเสมือนเป็นแกนโมเลกุลของดีเอ็นเอพันรอบแกนโปรตีนฮิสโตน 1 ¾ รอบ ยาวประมาณ 150 คู่เบส และมีโปรตีนฮิสโตน H1 จำนวน 1 โมเลกุลเชื่อมระหว่างแกนฮิสโตนและดีเอ็นเอที่พันรอบแกนฮิสโตน เรียกโครงสร้างนี้ว่า นิวคลีโอโซม (Nucleosome) ซึ่งมีเส้นผ่านศูนย์กลางยาว 110 Å หรือ 11 นาโนเมตร ดีเอ็นเอส่วนที่เหลือจะรวมกับโปรตีนฮิสโตนสร้างนิวคลีโอโซมอันใหม่ต่อไปเรื่อยๆ จนหมดความยาวของดีเอ็นเอ ทำให้เป็นสายโพลีนิวคลีโอโซม (Polynucleosome) ดีเอ็นเอที่อยู่ระหว่างนิวคลีโอโซมกับนิวคลีโอโซมเรียกว่า ดีเอ็นเอเชื่อม (Linker DNA) เมื่อดูจากกล้องจุลทรรศน์อิเล็กตรอนสายพอลีนิวคลีโอโซมมีรูปร่างคล้ายลูกปัดหรือลูกประคำซึ่งเชื่อมต่อเป็นสาย นิวคลีโอโซมและดีเอ็นเอที่เชื่อมมีชื่อเรียกรวมกันว่า โครมาโทโซม (Chromatosome)
องค์ประกอบของโปรตีนฮิสโตน
โปรตีนฮิสโตน 5 ชนิดต่างก็ประกอบด้วยกรดอะมิโนทั้งชนิดและจำนวนที่แตกต่างกัน คือ
- โปรตีนฮิสโตน H1 ประกอบด้วยกรดอะมิโนซึ่งมีความยาว 213 ตัว ซึ่งเป็นไลซีนมาก
- โปรตีนฮิสโตน H2A ประกอบด้วยกรดอะมิโนซึ่งมีความยาว 129 ตัว ซึ่งเป็นไลซีนและอาร์จินีนมาก
- โปรตีนฮิสโตน H2B ประกอบด้วยกรดอะมิโนซึ่งมีความยาว 125 ตัว ซึ่งเป็นไลซีนปานกลาง
- โปรตีนฮิสโตน H3 ประกอบด้วยกรดอะมิโนซึ่งมีความยาว 135 ตัว ซึ่งเป็นอาร์จินีนมาก
- โปรตีนฮิสโตน H4 ประกอบด้วยกรดอะมิโนซึ่งมีความยาว 102 ตัว ซึ่งเป็นอาร์จินีนและไกลซีนมาก
การจัดเรียงตัวของสายพอลินิวคลีโอโซม
สายโครมาโทโซม หรือสายพอลินิวคลีโอโซมเป็นโครงสร้างระดับแรกจะถูกจัดให้มีการเรียงตัวเข้าสู่โครงสร้างขั้นที่ 2 โดยนิวคลีโอโซม 8 โมเลกุลเรียงต่อกันเป็นวงกลม โดยโปรตีนฮิสโตน H1 และดีเอ็นเอเชื่อมอยู่ด้านใน เส้นผ่านศูนย์กลางของโครงสร้างขั้นที่ 2 นี้ยาว 30 นาโนเมตร ดครงสร้างนี้บางครั้งเรียกว่า โซลีนอยด์ (Solenoid) โครงสร้างโซลีนอยด์จะเรียงต่อกันเช่นนี้ไปเรื่อยๆ จนหมดความยาวของสายพอลีนิวคลีโอโซม โซลีนอยด์ก็จะรวมกับโปรตีนชนิดอื่นที่ไม่ใช่ฮิสโตน เช่น โปรตีนสคาฟโฟลด์ (Scaffold protein) สร้างเป็นโครงสร้างวง (Loop) มีทั้งวงเล็กและวงใหญ่ โดยมีความยาวของเส้นผ่านศูนย์กลางเฉลี่ย 300 นาโนเมตร เป็นโครงสร้างระดับที่ 3 และจัดเรียงต่อเป้นวงกลมโดยตรงกลางกลวงและเรียงซ้อนต่อกันเป็นทรงกระบอก จัดเป็นโครงสร้างระดับที่ 4 และปรากฏเป็นแท่งโครโมโซม มีเส้นผ่านศูนย์กลางประมาณ 700 นาโนเมตร
การค้นพบบทบาทของโครโมโซม
หลังจากการตั้งทฤษฏีเซลล์แล้ว นักชีววิทยาได้ค้นพบรายละเอียดของโครงสร้างต่างๆ ภายในเซลล์มากขึ้น โดยเฉพาะอย่างยิ่งในช่วงที่กำลังมีการแบ่งเซลล์ พบว่าภายในนิวเคลียสมีเส้นใยขนาดเล็กๆ ขดกันแน่นจนดูคล้ายเป็นแท่งเรียกว่า โครโมโซม (Chromosome) ซึ่งจะมีการจำลองตัวเองและแบ่งแยกออกไปยังเซลล์จำนวนเท่าๆกัน ปรากฏการณ์นี้ทำให้นักชีววิทยาเริ่มเข้าใจว่าการแบ่งเซลล์ทำให้มีการแบ่งโครโมโซมไปยังเซลล์ที่เกิดใหม่ เซลล์ที่เกิดใหม่จึงมีโครโมโซมเหมือนกับเซลล์เดิม ดังนั้นโครโมโซมจึงมีบทบาทสำคัญในการสืบทอดลักษณะทางพันธุกรรมของสิ่งมีชีวิตให้ต่อเนื่องกันไปได้
ใน พ.ศ. 2443 นักวิทยาศาสตร์ 3 คน คือ Correns ชาวเยอรมัน De Vries ชาวเนเธอแลนด์ และ Tschermak ชาวออสเตรีย ได้เสนอกฏเกณฑ์การถ่ายทอดลักษณะทางพันธุกรรมโดยมีข้อสรุปตรงกันว่า “การสร้างเซลล์สืบพันธุ์ของสิ่งมีชีวิตนั้นจะมีการแยกคู่ของยีนที่เป็นแอลลีลกัน” ซึ่งผลงานนี้เป็นที่ยอมรับกันอย่างกว้างขวาง ใน พ.ศ. 2455 Walter S. Sutton นักชีววิทยาชาวอเมริกัน และ Theodor Boveri นักชีววิทยาชาวเยอรมัน ซึ่งศึกษาเกี่ยวกับโครโมโซมและการถ่ายทอดลักษณะทางพันธุกรรม ได้เสนอทฤษฎี Chromosome Theory of Heredity ซึ่งอธิบายว่าหน่วยพันธุกรรมของสิ่งมีชีวิตอยู่บนโครโมโซม ทั้งนี้เพราะจากการศึกษาการแบ่งเซลล์แบบไมโอซิสของเซลล์อัณฑะของตั๊กแตน พบว่าฮอมอโลกัสโครโมโซมจะแยกออกไปอยู่ต่างเซลล์กัน เช่นเดียวกับการแยกของยีนที่เป็นแอลลีลกันตามกฎแห่งการแยกตัว ทำให้ได้ข้อสรุปว่ายีนจะต้องอยู่บนโครโมโซม ดังนั้นโครโมโซมจึงมีบทบาทสำคัญในการถ่ายทอดลักษณะทางพันธุกรรมของสิ่งมีชีวิต
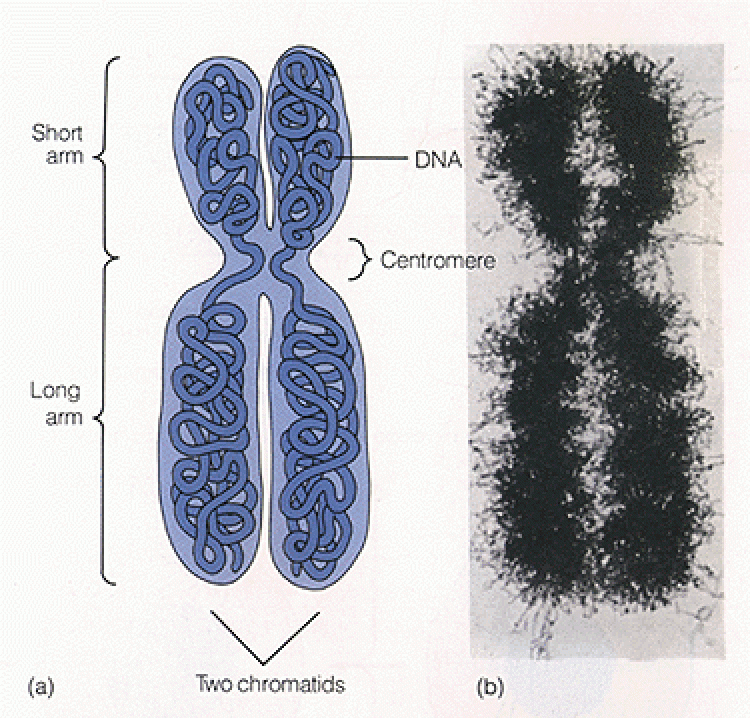
ส่วนสำคัญของโครโมโซม
โมเลกุลของดีเอ็นเอในนิวเคลียสที่เป็นเกลียวคู่รวมกับโปรตีนไม่ว่าจะเป็นโปรตีนฮิสโตน หรือโปรตีนที่ไม่ใช่ฮิสโตน เรียกว่า โครมาติน (Chromatin) บนแท่งโครโมโมโซม บางส่วนของโครมาตินขดตัวแน่นเรียกว่า เฮเทอดรโครมาติน (Heterochromatin) สามารถสังเกตเห็นภายใต้กล้องจุลทรรศน์เมื่อย้อมสีโครโมโซมด้วยสี Giemsa หรือสี Carmine เฮเทอโรโครมาตินจะติดสีเข้ม ส่วนของโครโมโซมที่เกิดจากโครโมโซมที่ขดตัวอย่างหลวมๆ เรียกว่า ยูโครมาติน (Euchromatin) จะติดสีจางกว่า โดยทั่วไปโครโมโซมมีรอยคอด (Constriction) ซึ่งเกิดจากโครโมโซมที่ขดตัวแน่นมาก 1 แห่ง เรียกว่า เซนโทรเมียร์ (Centromere) หรือไพรมารีคอนสตริกชัน (Primary constriction) แต่โครโมโซมบางแท่งมีรอยคอด 2 แห่ง รอยคอดที่ 2 เรียกว่า เซคันดารีคอนสตริกชัน (Secondary constriction) โครโมโซมที่มีรอยคอด 2 แห่งเรียกว่า โครโมโซมซาเทลไลท์ (Satellite chromosome) รอยคอดที่ 1 หรือเซนโทรเมียร์นั้นมีความสำคัญต่อการจำแนกชนิดของโครโมโซมและการเคลื่อนที่ของโครโมโซมขณะที่มีการแบ่งเซลล์ ส่วนรอยคอดที่ 2 ของโครโมโซมซาเทลไลท์บางแท่งทำหน้าที่สร้างนิวคลีโอลัส บริเวณนี้จึงเรียกว่า บริเวณนิวคลีโอลาร์ออร์กาไนเซอร์ (Nucleolar organizer region)
กลับไปที่เนื้อหา
รูปร่าง ขนาด และจำนวนโครโมโซม
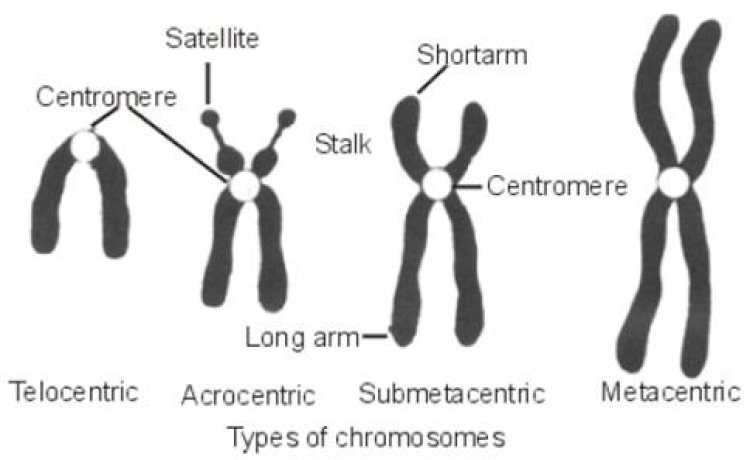
โครโมโซมเป็นโครงสร้างที่อยู่ในนิวเคลียสของเซลล์ ในภาวะปกติจะมองไม่เห็นโครโมโซม เพราะมีสภาพเป็นเส้นใยขนาดเล็กๆ สานกันอยู่ภายในนิวเคลียส เมื่อเข้าสู่ระยะการแบ่งเซลล์จะมีการจำลองเส้นใยโครมาทินขึ้นมาเป็นเส้นคู่ที่เหมือนกัน และมีการขดตัวพันกันจนมองเห็นเป็น 2 แท่งติดกัน ซึ่งจะเห็นชัดเจนที่สุดในระยะเมทาเฟส ระยะนี้โครโมโซม 1 โครโมโซมจะประกอบด้วย 2 โครมาทิด (Chromatid) ซึ่งติดกันอยู่ที่เซนโทรเมียร์ (Centromere) จากตำแหน่งของเซนโทรเมียร์มีผลทำให้โครโมโซมมีรูปร่างแตกต่างกัน 4 แบบ คือ
- โครโมโซมแบบเทโลเซนทริก (Telocentric chromosome) เป็นโครโมโซมที่มีเซนโทรเมียร์อยู่ตรปลายสุด ทำให้มองเห็นโครโมโซมเป็นแท่งหรือแขนเพียงข้างเดียว
- โครโมโซมแบบอะโครเซนทริก (Acrocentric chromosome) เป็นโครโมโซมที่มีเซนโทรเมียร์อยู่ค่อนข้างไปทางส่วนปลายด้านใดด้านหนึ่ง ทำให้มองเห็นโครโมโซมเป็นแท่งเช่นกัน แต่มีแขน 2 ข้างยาวแตกต่างกันมาก
- โครโมโซมแบบซับเมทาเซนทริก (Submetacentric chromosome) เป็นโครโมโซมที่มีเซนโทรเมียร์ไม่อยู่ตรงกลาง ทำให้แขน 2 ข้างยาวไม่เท่ากัน มองเห็นคล้ายรูปตัวเจ (J)
- โครโมโซมแบบเมทาเซนทริก (Metacentric chromosome) เป็นโครโมโซมที่มีเซนโทรเมียร์อยู่ตรงกลางพอดี ทำให้แขน 2 ข้างยาวเท่ากัน มองเห็นคล้ายรูปตัววี (V)
ขนาดของโครโมโซมในสิ่งมีชีวิตแต่ละชนิดมีความแตกต่างกัน และมีการผันแปรตามระยะต่างๆ ของการแบ่งเซลล์ (โครโมโซมมีขนาดสั้นที่สุดในระยะเมทาเฟส) ขนาดของโครโมโซมไม่เป็นสัดส่วนกับจำนวนยีนบนโครโมโซม คือ โครโมโซมที่มีขนาดความยาวมากอาจมีจำนวนยีนอยู่น้อยก็ได้ แต่ขนาดของโครโมโซมอาจมีความเกี่ยวข้องสัมพันธ์กับปัจจัยบางอย่างดังต่อไปนี้
- สารโคลซิซิน (Colchicine) เป็นสารยับยั้งการสร้าง Spindle fiber ทำให้โครโมโซมหยุดอยู่ที่ระยะเมทาเฟส ซึ่งมีขนาดสั้นที่สุด
- สารที่ใช้ทำ Pretreatment เช่น α – bromonap thalene ทำให้โครโมโซมหดตัว เห็นโครโมโซมมีขนาดสั้น
- สารที่จำเป็นต่อการสังเคราะห์โครมาทิน พบว่าเซลล์ที่มีการแบ่งเซลล์บ่อยๆ จะมีขนาดโครโมโซมเล็กกว่าเซลล์ที่มีระยะอินเตอร์เฟสค่อนข้างยาว
- สารอาหารที่ใช้สำหรับเพาะเลี้ยงเซลล์ จากการศึกษาพบว่าโครโมโซมของเซลล์ที่เลี้ยงอาหารซึ่งมีฟอสเฟตความเข้มข้นสูง จะมีขนาดใหญ่กว่าโครโมโซมของเซลล์ที่เลี้ยงในน้ำหรือมีฟอสเฟตน้อยๆ
- เซลล์ที่มีการแบ่งตัวในอุณหเซลล์ที่มีการแบ่งตัวในอุณหภูมิต่ำ จะมีการหดตัวของโครโมโซมมากกว่าเซลล์ที่แบ่งตัวในอุณหภูมิสูงๆ
จำนวนโครโมโซมของสิ่งมีชีวิตโดยทั่วไปจะแตกต่างกัน ในเซลล์ร่างกาย (Somatic cell) จะมีโครโมโซม 2 ชุด หรือเรียกว่ามีจำนวนโครโมโซมเป็น 2n หรือดิพลอยด์ (Diploid) ส่วนเซลล์สืบพันธุ์ (Sex cell) จะมีโครโมโซมเพียงชุดเดียวหรือเรียกว่ามีโครโมโซม n หรือแฮพลอยด์ (Haploid) ในภาวะปกติสิ่งมีชีวิตแต่ละชนิดจะมีจำนวนโครโมโซมคงที่
กลับไปที่เนื้อหา

 (400744)
(400744)  (355326)
(355326)  (90577)
(90577) 